

Botanik online 1996-2004. Die Seiten werden nicht mehr bearbeitet, sie bleiben als historisches Dokument der botanischen Wissenschaft online erhalten!
Epidermisabdrucke, hergestellt durch Auftragen eines Acrylklebers (Acrifix®) auf die Blattoberfläche. Nach dem Erhärten wurde der Kunststoffilm einfach abgezogen und in einem Interferenzkontrastmikroskop mikroskopiert. Links: Blattoberseite einer Palme (Hovea belmoreana) mit extrem kleinen, in Reihen angeordneten (für Monokotyledonen typischen) Epidermiszellen. Rechts: Blattunterseite einer Cracculaceae (Aeonium acuneatum) mit Spaltöffnungspaaren und extrem großen Epidermiszellen.
Es gibt zwei Techniken, um sich ein anschauliches Bild des strukturellen Aufbaus von Epidermisoberflächen zu machen. Einmal die Rasterelektronenmikroskopie, zum anderen die mikroskopische Analyse von Kunststoffabdrucken. Der Erfolg der beiden Techniken wird durch eine Anzahl von Beispielen demonstriert (blaue Textbereiche anklicken!).
Die "eigentlichen", d.h., die am wenigsten spezialisierten Epidermiszellen machen die Hauptmasse der Zellen des Abschlußgewebes aus. Sie sind in der Aufsicht entweder polygonal (von platten- oder tafelförmiger Gestalt) oder gestreckt. Die zwischen ihnen ausgebildeten Wände sind vielfach gewellt oder gebuchtet. Wodurch diese Form während der Entwicklung induziert wird, ist unbekannt, die vorliegenden Hypothesen erklären den Sachverhalt nur unbefriedigend. Gestreckte Epidermiszellen findet man an Organen oder Organteilen, die selbst gestreckt sind, so z.B. an Stengeln, Blattstielen und Blattrippen sowie an den Blättern der meisten Monokotyledonen. Ober- und Unterseite von Blattspreiten können von unterschiedlich strukturierten Epidermen bedeckt sein wobei sowohl die Form der Zellen, die Dicke der Wände als auch die Verteilung und Zahl spezialisierter Zellen (Stomata und/oder Trichome) pro Flächeneinheit variieren kann. Große Variationen findet man auch innerhalb einzelner Familien, z. B. bei den Crassulaceen.
Oft ist die Außenwand der Zellen dicker als die der übrigen Wände. Besonders deutlich tritt das bei der Epidermis von Koniferennadeln, derben Blättern vieler Arten des Regenwaldes und bei Xerophyten (Pflanzen trockener Standorte) in Erscheinung. Dünne Wände findet man bei Wasserpflanzen. In vielen Samen verstärkt sich die Wand im Verlauf der Reifung und erfüllt fast das gesamte Lumen der Zelle, der Protoplast wird verdrängt und degeneriert. Die Epidermiszellen der meisten Arten sind chloroplastenfrei. Ausnahmen findet man bei Farnen, Wasser- und einigen Schattenpflanzen.
Meist ist die Epidermis einschichtig, jedoch sind bei Arten aus mehreren Familien (Moraceae: hier die meisten Ficus-Arten, Piperaceae: Peperonia [Peperonie], Begoniaceae, Malvaceae u.a.) mehrschichtige wasserspeichernde Epidermen nachgewiesen worden , die durch perikline Teilungen aus einer ursprünglich einschichtigen Gewebelage hervorgegangen sind. Epidermiszellen sondern nach außen eine Cutinschicht (Kutikula) ab, die als ein ununterbrochener Film alle epidermalen Oberflächen überzieht. Sie kann entweder glatt oder durch Vorwölbungen, Leisten, Falten und Furchen strukturiert sein.
Doch nicht immer beruht eine durch Betrachtung der Oberfläche sichtbare Faltung der Kutikula auf der Ausbildung von Kutikularleisten. Es gibt durchaus Fälle, wo eine Kutikulafaltung nur der Ausdruck der darunterliegenden Ausstülpungen der Zellwand ist. Zum Nachweis solcher Strukturen ist man auf die Auswertung von Querschnitten durch die Kutikula und die darunterliegenden Epidermiszellen angewiesen.
In einigen Fällen, so bei der Frucht der Tomate, ist die Kutikula durch eingelagerte Carotinoide pigmentiert. Zusätzlich werden vielfach Wachse, Öle, Harze, Salzkristalle und wasserlösliche (hydrophile) Schleime abgesondert. Besonders häufig tritt eine Verschleimung bei der Samenbildung in Erscheinung. Durch Wachsausscheidungen wird die Benetzbarkeit der Blätter, mehr noch als durch die Kutikula selbst, verhindert. Vielfach sind diese Wachse auch in sich strukturiert . Starker Wachsbelag verleiht pflanzlichen Oberflächen ein weißliches Aussehen. Er wirkt auf zweierlei Weise. Einmal schränkt er die Transpiration (den Wasserverlust) ein, zum anderen bewirkt er (aufgrund der hellen Färbung) eine Reflexion des Lichts und schützt die Pflanze damit vor übermäßiger Erhitzung. Die aufgelagerten epicuticularen Wachse verursachen zudem Mikroturbulenzen und verringern somit die Adh�sion von Wasser und partikelf�rmigen Kontaminationen
Die Unbenetzbarkeit von Blattoberfächen ist lange bekannt und gut untersucht. Es wurde jedoch weitgehend übersehen, daß unbenetzbare Oberfächen auch nahezu unverschmutzbar sind. Dieser Zusammenhang wurde erst in jüngster Zeit detailliert untersucht und experimentell belegt. Da er sich besonders gut an den großen schildförmigen Blättern der heiligen Lotuspflanze (Nelumbo nucifera) demonstrieren läßt, dem Symbol für Reinheit in asiatischen Religionen, wurde er von W. BARTHLOTT und C. NEINHUIS "Lotus-Effekt" genannt.
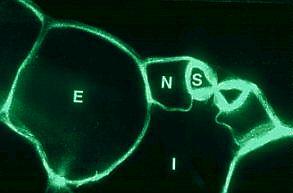
Querschnitt durch eine Spaltöffnung von Rhoeo discolor. S Schließzelle, N Nebenzelle, E nicht spezialisierte Epidermiszelle I Interzellularraum. (R. KAPPLER, 1984)
Eine wichtige Funktion der Epidermis wird von den Spaltöffnungen, den Stomata, wahrgenommen. Die vollständige funktionelle Einheit wird als Stomatakomplex oder Spaltöffnungsapparat bezeichnet. Dazu gehören zwei chloroplastenhaltige Schließzellen, zwischen denen eine Pore (ein Spalt) vorhanden ist, sowie zwei bis vier benachbarte chloroplastenfreie Nebenzellen. In Querschnitten wird deutlich, daß die Schließzellen unterschiedlich dicke Zellwände besitzen und ihnen ein Interzellularraum (eine "Atemhöhle") unterlagert ist, der mit den übrigen Interzellularen des betreffenden pflanzlichen Gewebes kommunizierend verbunden ist. Nach Bedarf kann der Spalt erweitert oder geschlossen werden. Die Spaltöffnungen regeln somit Transpiration (Wasserabgabe) und Kohlendioxyd-Aufnahme. Sowohl der Wassergehalt als auch die Kohlendioxyd-Konzentration im pflanzlichen Gewebe wirken als Regulatoren des Öffnungszustands. Durch Gestaltveränderung regulieren die Schließzellen die Porengröße.
Stomata kommen an allen oberirdischen Pflanzenteilen vor. Ihre Anzahl liegt in der Größenordnung von 100 - 300 / Quadratmillimeter, kann aber - artspezifisch - auch in viel weiteren Grenzen schwanken., ebenso variabel können auch die Verteilung und die Spaltendurchmesser sein. An Wurzeln, in der Epidermis der chlorophyllfreien Sprosse der parasitischen Arten Monotropa hypopitys (Fichtenspargel) und Neottia nidus-avis (Nestwurz) sowie bei einigen submers lebenden Wasserpflanzen fehlen sie, bei anderen Wasserpflanzen hingegen sind sie normal ausgeprägt. Sie kommen auch in gefärbten und weißen Blütenblättern vor, doch sind sie hier oft funktionslos. Bei den parallelnervigen Blättern der meisten Monokotyledonen, einigen Dikotyledonen sowie in den Nadeln der Koniferen sind sie in parallel zueinanderliegenden Reihen angeordnet.
Ihre Entstehung geht auf Schließzellenmutterzellen zurück, die wiederum in der Epidermis in regelmäßigen Abständen durch inäquale Teilungen angelegt werden. Die Initiale der Spaltöffnung ist dabei die kleinere, plasmareichere der beiden Tochterzellen. Die beiden Schließzellen entstehen aus ihr durch eine äquale Teilung, die Bildung der Nebenzellen kann auf ganz unterschiedliche Weise erfolgen. Die Entwicklung der Stomata in einem Blatt erfolgt meist asynchron. In parallelnervigen Blättern folgt ihre Entwicklung der sukzessiven Differenzierung der einzelnen Blattabschnitte. Die Differenzierungswelle verläuft basipetal, d.h., sie beginnt an der Blattspitze und dehnt sich in Richtung der Blattbasis aus.
Bei netznervigen Blättern sind verschiedene Entwicklungsstadien der Stomata mosaikartig über die gesamte Blattfläche verteilt.
Variationen zum Thema: Da es unterschiedliche Entstehungswege für Spaltöffnungsapparate gibt, ist zu erwarten, daß auch morphologische Varianten vorkommen. Als typische Ausnahmeform werden immer die Schließzellen von Gramineen genannt, die bei vielen Arten hantelförmig strukturiert sind. Von den Spaltöffnungen ableitbar sind die Hydathoden (Wasserspalten), die oft an den Enden der Leitungsbahnen zu finden sind. Die Schließzellen sehen zwar noch so wie bei den Spaltöffnungen aus, lassen sich aber nicht mehr schließen. Eine Wasserabscheidung durch sie wird als Guttation bezeichnet. Charakteristisch gebaute Hydathoden kommen u.a. an Blatträndern der Kapuzinerkresse (Tropaeolum majus), des Frauenmantels (Alchemilla vulgaris: s.nebenstehende Abbildung. Aus. J. v. SACHS: "Vorlesungen über Pflanzen-Physiologie", 1877) und an den Blattspitzen vieler Gräser vor. Im Guttationswasser gelöste Salze, sowie Zucker und andere organische Substanzen kristallisieren nach Verdunstung des Wassers an der Austrittsstelle aus. Als typische Beispiele hierfür können die Kalkabsonderungen der Steinbrecharten (Saxifraga) und die Salzdrüsen der Halophyten (Salzpflanzen) genannt werden. |
 |
Epidermale Anhangsgebilde verschiedener Form, Struktur und Funktion werden als Trichome bezeichnet. Sie treten als Schutz-, Stütz- und Drüsenhaare in Form von Schuppen, verschiedenen Papillen und bei Wurzeln als absorbierende Haare auf. An ihrer Bildung sind allein Epidermiszellen beteiligt. Oft entsteht ein Trichom aus nur einer solchen Zelle, manchmal sind an der Entstehung mehrere beteiligt.
Die Trichome sind von den Emergenzen (z.B. den Stacheln) und den Kurztrieben (z.B. den Dornen) zu unterscheiden, weil jene nicht nur Zellen der Epidermis, sondern auch Zellen anderer Gewebe mit enthalten.
Haare: An pflanzlichen Oberflächen kommen Haare in mannigfaltiger Ausbildung vor. Sie können ein- oder vielzellig, verzweigt oder unverzweigt, lebend oder tot sein. Ihre Wand kann durch Silikat, Calciumcarbonat oder andere Einlagerungen verhärtet sein und ihnen damit einen borstenartigen Charakter verleihen. Solche steifen Borsten (z.B. bei den Boraginaceae und den Cruciferae) schützen die Pflanzen vor Tierfraß. Viele, vor allem die stark verzweigten Haare dienen dem Transpirationsschutz. Es ist bekannt, daß Pflanzen trockener Standorte entweder sukkulent sind (und damit auch eine dicke Kutikula besitzen) oder über einen dichten, silbrig erscheinenden Haarfilz verfügen. Bei Stachys lanata (Filziger Ziest) wurden 120 Haare / Quadratmillimeter gezählt. Eine mikroskopische Analyse zeigt, daß Haarzellen oft stark verzweigt und abgestorben sind. Daraus ergeben sich für die Pflanze gleich drei Vorteile:
Das Lumen toter Zellen ist lufterfüllt. Es verleiht den Zellen ein weißlich-silbriges Aussehen. Ein großer Teil des eingestrahlten Lichts wird reflektiert. Der Effekt ist der gleiche, der mit dicken Wachsschichten erreicht wird.
Über der Blattoberfläche wird ein zirkulationsberuhigter Raum geschaffen. Mit anderen Worten: Wasserverluste an der Blattoberfläche werden auf ein Minimum reduziert.
Durch das Absterben der Zellen wird die wasserverlierende Oberfläche drastisch reduziert. Lebten die Zellen, wäre wegen der durch die Verzweigungen bedingten großen Oberfläche ein zu starker Wasserverlust unvermeidlich.
Haare sind an Blattoberflächen regelmäßig angeordnet,
sie werden stets in annähernd gleichen Abständen voneinander
angelegt. Vergleichbare Muster treten auch bei den Stomata auf. Damit stellt
sich für uns die Frage nach den Ursachen dieser Regelmäßigkeit.
Zwar können wir heute darauf noch keine abschließende Antwort
geben, wohl aber ein mathematisches Modell präsentieren, das nach
Regeln aufgestellt wurde, die auch in den pflanzlichen Geweben gelten dürften.

Aerenchym mit "inneren Haaren", die eine Verholzungsreaktion zeigen (Nymphaea spec.): Handschnitt durch den Blattstiel. Färbung mit Phloroglucin-HCl. (Aufn.: W. KASPRIK).

Ein Kaleidoskop verschiedener Haare: Ausgesprochen lange (1-6 cm), einzellige und unverzweigte Haare, deren Wand aus nahezu reiner Cellulose besteht, umgeben die Samen von Gossypium (Baumwolle). Verzweigt sind sie z.B. bei Lobelia bei Arabis alpina und bei Malacantha alnifolia (nebenstehendes Bild). Vielzellige Haare können aus einer oder aus mehreren Zellreihen bestehen. Typische Beispiele sind die Etagenhaare der Platane (Platanus hybrida) oder die der Königskerze (Verbascum nigrum). Haare auf Blättern der Eiche (Quercus robur) sehen büschelförmig, die vieler Malvaceae sternförmig aus. Bei der Ölweide (Elaeagnus angustifolia) sind sie schirmartig gebaut (Schuppenhaare). Bei den Bromeliaceen haben derartige Schuppen die Funktion von Absorptionshaaren. Die Feuchtigkeit der Atmosphäre wird durch Kapillarkräfte gesammelt und kommt der Pflanze zugute. Außer Haaren auf Oberflächen kommen auch sogenannte "innere Haare" vor, z. B. bei der Seerose
Drüsenhaare bestehen aus einem mehrzelligen Stiel und einem ein- oder vielzelligen Köpfchen. Beispiele für die Entwicklung der Haare: Origanum (Lamiaceae), Adenocaulon (Compositae), Citrus deliciosa (Rutaceae). Drüsenhaare auf Blättern des Tabaks (Nicotiana tabacum) haben vielzellige Köpfchen, bei Primeln (z.B. Primula sinensis) und Pelargonien (z.B. Pelargonium zonale) sind sie einzellig. Das Sekret (von Pelargonium zonale) ist ein ätherisches Öl. Die Drüsenzellen selbst sind plasmareich, die Sekretion erfolgt durch die Zellwand. Das Öl sammelt sich an der Zelloberfläche und erscheint unter dem Mikroskop daher als stark lichtbrechende Kappe, die von einem dünnen Film aus Kutikulaanteilen und Zellwandbestandteilen bedeckt ist. Nach deren Platzen wird das Sekret freigesetzt. In der Fruchtschale, z.B. der Citrusfrüchte sind eingesenkte sekretorische Kavernen (Drüsen) enthalten.
Die Staubfäden (Filamente) von Tradescantia virginiana sind
von mehrzelligen Haaren umgeben, die einzelne Zelle sieht tonnenförmig
aus. Der Protoplast ist wandständig, der Vakuoleninhalt durch einen
rotvioletten Farbstoff (ein Anthocyan) gefärbt. Die Vakuole ist von
zahlreichen, unterschiedlich dicken Plasmasträngen durchsetzt. Der
Kern liegt zentral, und es sieht so aus, als sei er an den Plasmasträngen
aufgehängt. Da die Plasmastränge ihre Form ständig ändern,
wird der Kern, wie an elastischen Gummibändern hängend, ständig
hin- und hergezogen. In den Plasmasträngen strömt das Plasma
lebhaft und transportiert deutlich erkennbare Granula. Die Strömung
ist streng gerichtet, wobei in vielen Strängen ein Gegenverkehr auf
getrennten Bahnen sichtbar ist. Wegen der leichten Erkennbarkeit und Regelmäßigkeit
der Strömung sind die Staubfadenhaare der Tradescantia virginiana
zu einem klassischen Demonstrationsobjekt für diese Zellaktivitäten
geworden.
Aus einem anderen Grund wurden die Staubfadenhaare der Flockenblume (Centaurea jacea) und der Kornblume (Centaurea cyanus) bekannt. Sie sind berührungsempfindlich und lösen eine Reizbewegung der Staubfäden aus.
Brennhaare der Brennessel (z.B. Urtica dioica): Ein Brennhaar ist eigentlich eine Emergenz, es ist zweiteilig und besteht aus einem vielzelligen Sockel, an desssen Bildung auch subepidermal gelegene Zellen beteiligt sind, und einer in ihn eingelassenen Haarzelle. Der untere (basale) Abschnitt der Haarzelle wird Bulbus genannt. Er ist von den Sockelzellen becherfömig umgeben. An seinem oberen Ende läuft das Haar spitz aus und endet in einem seitlich angesetzten Köpfchen. An der Übergangsstelle ist die Zellwand merklich dünner als in den übrigen Abschnitten. Eingelagerte Silikate machen sie zudem spröde, so daß das Köpfchen bei Berührung leicht abbricht und an der Sollbruchstelle eine Spitze hinterläßt, die einer Einstichkanüle gleicht. Wegen der Starrheit der Wand wird der Druck ungepuffert auf den Bulbus übertragen, dessen Inhalt (Natriumformiat, Acetylcholin und Histamin) durch die Kanüle ausgepreßt und damit gegebenenfalls in eine Wundstelle injiziert wird.
Weitere Trichome und Spezialfunktionen der Epidermis. Papillen sind Ausstülpungen der Epidermisoberfläche. Das Lehrbuchbeispiel hierfür sind die Papillen auf Blütenoberflächen des Stiefmütterchens (Viola tricolor) sowie die Blattoberflächen vieler Arten im tropischen Regenwald. Sie verleihen der Oberfläche eine samtartige Konsistenz. Einige Zellen von Epidermen können als Wasserspeicher ausgebildet sein. Ein typisches Beispiel stellen die Blasenzellen an Oberflächen vieler Mittagsblumenarten und anderer Sukkulenten dar. Bei manchen Pflanzen, z.B. bei der Glockenblume (Campanula persicifolia) sind die Außenwände der Epidermis linsenförmig verdickt. Sie wirken damit wie Sammellinsen und bündeln das Licht, das seinerseits von spezifischen Rezeptoren (Lichtperzeptoren) wahrgenommen und in eine physiologische Reaktion umgesetzt wird.

Blasenzelle in Aufsicht. Epidermisabdruck.
Monanthes lowei. (Präparat J. THIEDE)