STEMS II
Woody Dicot Vine - Aristolochia (Dutchman's pipe).


Vines like Aristolochia commonly have wide interfascicular areas in the primary state of growth and correspondingly wide rays in the secondary body.
Examine slides of four stages of development
(youngest = A, oldest = D).
Also study
slides prepared from living stems of Aristolochia.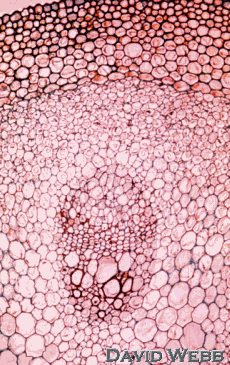 This genus is largely native to North and South America but is also
found in the Mediterranean.
This genus is largely native to North and South America but is also
found in the Mediterranean.
The youngest stems (slide A) are still undergoing primary growth. The epidermis is the surface layer. Just beneath this there are 10-15 layers of collenchyma and parenchyma cells that form the outer cortex. The innermost of these cell layers is slightly larger than the others and constitutes a starch sheath.
Inside this are 10-15 layers
of cells with smaller diameters. These will develop
into perivascular fibers. Perivascular means surrounds the vascular
system. Inside this one sees immature vascular bundles.
develop
into perivascular fibers. Perivascular means surrounds the vascular
system. Inside this one sees immature vascular bundles.
The bundles (fascicles) are separated by parenchyma that comprises the interfascicular areas.
The center of the stem is pith composed of large parenchyma cells.
 |
 |
 |
| Young Stem stained with Toluidine Blue | Vascular Bundle stained with Toluidine Blue | Stem stained with Phloroglucinol & viewed with crossed polarizers |
Locate these major structures in commercial slides and fresh sections.
Stain with Phloroglucinol and Toluidine Blue.
Don’t forget to use your polarizers.
One-year-old stems (slide B) have completed primary growth. Tissues barely evident in the youngest sections have now matured.
The vascular cambium should be prominent between the primary xylem and phloem, and the interfascicular cambium is just forming.
Locate the major structures and regions in commercial slides
 |
 |
 |
| Vascular Bundle & interfascicular zone | Active Vascular Cambium | Induction of cambial activity in the interfascicular region |
Two- three year old stems (slide C) exhibit considerable secondary growth.
 Vascular Bundles in a 3 year-old stem |
 Pith in a 3 year-old stem |
The Vascular Cambium and the Cork Cambium (phellogen) produce secondary growth. These produce internal pressures due to the production and enlargement of the new cells they produce.
During enlargement the ring of perivascular (extraxylary) fibers often becomes broken. The gaps in the ring are filled with parenchyma cells, some of which develop into thick-walled sclereids. In this case, pressure has induced meristematic activity in cells.
 |
 |
| Results of secondary growth on the stem fibers | Effects of secondary growth on the phloem |
The epidermis is still intact except in a few places where lenticels are forming from the phellogen. Since the interfascicular cambium is composed only of ray initials, the vascular bundles remain distinct, and are separated by wide rays.
The xylem shows two growth layers.
Many pith cells are crushed, as a consequence both of movement of the vine and of inward pressure of developing xylem tissue.
The fact that many vines have isolated vascular bundles or wide areas of parenchyma in the xylem suggests that this might have functional significance. Considering the manner in which vines grow, can you imagine what function this tissue organization may have?
Older stems (slide D) clearly show periderm formation. In Aristolochia the periderm develops first in isolated vertical strips. Thus, as seen in cross sections, parts of the stem's circumference have an intact epidermis while other parts have a well developed periderm with several layers of cork cells (phellem), cork cambium (phellogen) and a relatively wide phelloderm.
The
Phellogen (Cork Cambium) produces the Phellem (Cork) to the
outside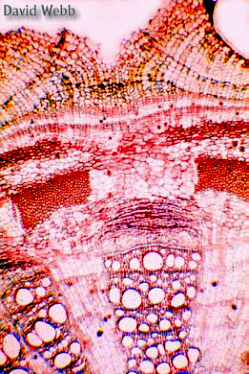 and the Phellogen towards the Inside.
and the Phellogen towards the Inside.
The Phellogen can be located by following the two files of vacuolate cells towards the centrally located meristematic cells.
Remember that meristematic cells are densely cytoplasmic and lack prominent vacuoles.
The phelloderm is usually not prominent and may consist of a single layer of parenchyma.
Note the lenticels in the periderm. These are areas of hyperactive cell division compared to the rest of the Phellogen.
The xylem shows several growth layers. As the individual wedges of vascular tissues increase in size, new rays develop within them.The pith and some of the inner parts of the interfascicular areas are almost completely crushed.
Coleus Stems
The development
of secondary growth in stems  is
readily seen by making a series of cross sections from a Coleus
stem.The large primary bundles are conveniently located in the corners of
its square stem. By taking sections from more mature parts of the stem
the pattern of development is readily discerned!Examine the series of photos
below, then observe fresh sections.
is
readily seen by making a series of cross sections from a Coleus
stem.The large primary bundles are conveniently located in the corners of
its square stem. By taking sections from more mature parts of the stem
the pattern of development is readily discerned!Examine the series of photos
below, then observe fresh sections.
| Developmental Sequence of Vascular Differentiation in Coleus | |
| Coleus and other mints (Lamiaceae) have large primary vascular bundles occur in each corner of the stem. The bundles have Bast Fibers, Phloem and Xylem. A Vascular Cambium is present between the xylem and phloem. It divides periclinally to produce files of cells. Cells displaced towards the interior become xylem. Those displaced towards the periphery become phloem. Cambial activity spreads to the interfascicular parenchyma. This spreads laterally until a complete ring of cambium encompasses the stem and unites the bundles. |
|
Study a series fresh sections of Coleus or another related plant stained with Phloroglucinol or Toluidine Blue.
Be sure to use your polarizers.
Compare
Growth from the vascular cambium increases the girth or diameter of the stem. The vascular cambium of Coleus usually produces more xylem than phloem. The thick walled xylem cells accumulate like loaves of bread which enlarge as they mature and have a hard crust after they are baked. These files of sturdy cells create pressure towards the inside and outside of the plant. The thin walled cells of the pith and phloem are crushed as the pressure increases. The vascular cambium replenishes the phloem. However, the amount of active phloem may be limited to a small zone. Fibers may be present in the secondary phloem. These protect the sieve elements and allow for the expansion of the active phloem.
The Phellogen (Cork Cambium) is another lateral meristem. It usually develops from subepidermal cells in stems. It divides periclinally to produce rows of cork (Phellem) cells towards the exterior. Cork cells are dead at maturity and have suberized walls.
 |
 |
 |
| Early stages in Periderm Development | ||
Suberin is a lot like cutin. It protects the stem because It is impervious to water and most pathogens, and provides some insulation. It stains positively with Phloroglucinol.
Look
Study unstained sections
Deploy your Polarizers!!!!!
Stain


What accounts for the Layers that you see in the Periderm?

These questions are really getting to be a major annoyance! Can't a guy or gal enjoy some microscopy without all of these nagging questions!
Optional
We can study the transition from primary to secondary growth by studying Cotton (Gossypium tomentosm) and/or Coffee (Coffea arabica).
Hawaiian cotton is indigenous to these islands but is probably part of a larger taxon that is widely distributed in the Pacific. It does not produce the finest fibers but it has important traits that have been used in cotton breeding programs. This supports the concept of population/species preservation in that valuable genes may exist in natural populations. If wild populations are destroyed, the beneficial genes are also destroyed.
Locate the youngest internodes on stems of either species.
Study cross sections and stain with Phloroglucinol and Toluidine blue.
Examine more mature internodes which have a brown exterior and treat as above.
Locate the following:
Dermal Layer (Epidermis – Periderm (Phellogen [Cork Cambium] – Phellem [Cork])
Cortex (Collenchyma – Sclerenchyma – Parenchyma)
Vascular Tissue (Phloem Fibers – Phloem – Vascular Cambium – Xylem
Interfascicular Region (Parenchyma – Vascular Cambium)
Pith (Parenchyma – Other Cell Types)
Study
Primary xylem will jut into the pith due to the production of secondary xylem. Secondary xylem is usually more linear in its organization and may have a distinctly different cell composition compared to primary xylem.
The phloem fibers in the cortex can also mark the location of primary vascular bundles.
Woody dicot Tilia (basswood) stem.
 Tilia sp. |
 Tilia flowers |
 Tilia fruits |
Tilia
 studied
for the stem anatomy of woody angiosperms. Its characteristics
encompass a good sample of anatomical features for this kind of plant.
studied
for the stem anatomy of woody angiosperms. Its characteristics
encompass a good sample of anatomical features for this kind of plant.
We will study secondary xylem and phloem in depth later in the course. Do not worry about the details. My goal is to simply introduce the anatomical basics for these secondary tissues.
Study commercial
slides of stems in several 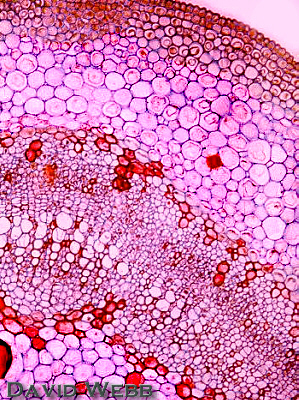 stages of
development.
stages of
development.
Work from the outside towards the inside of the stem!!!
The following details may be observed:
In younger stages the epidermis is present.
In older
stems it is replaced by a periderm containing
several layers of tannin-filled cork cells. Tannins are brown and
impart a dark color to the Cork. The smallest periderm cells that
contain protoplasts with nuclei are the innermost cells in each
radial file of cork cells. These cells are the phellogen or cork
cambium.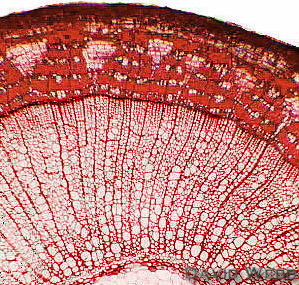
The Cortex is composed of collenchyma and parenchyma. The large cells with red-staining contents are mucilage cells.
The Vascular Tissues form a continuous ring.
The Phloem
in older stems appears to be divided into two kinds of wedges
with opposite orientations.
The wedge that points towards the outside
contains sieve-tube members, companion cells, phloem parenchyma and phloem fibers.
The fibers occur in tangential bands (parallel to the surface) alternating with bands that contain the rest of the phloem components.
The outermost fiber bundles are the primary phloem fibers, the others arose from the vascular cambium and are part of the secondary phloem.
The other Wedge is principally composed of
parenchyma. These are dilated phloem rays. Dilated means broadened or expanded.
In addition to the dilated rays, there are narrow undilated rays.
Note the continuity of the rays from the phloem to the xylem.
The stems in slides labeled "young stem" are just starting to form secondary xylem.
In older
sections from one to several increments of secondary xylem occur, and the primary xylem is next to the pith.
occur, and the primary xylem is next to the pith.
The secondary xylem of Tilia contains vessel elements (widest cells), tracheids, fibers (narrow cells with relatively thick walls), and xylem parenchyma (small dark cells).
The Pith contains thick-walled cells with dark globules of tannin.
The large
"spaces" in the pith, which may be filled with red-staining  material, are mucilage ducts.
material, are mucilage ducts.
Woody Gymnosperms – Araucaria, Podocarpus or
Pinus stems.
 Pinus strobus |
 Araucaria sp. |
The initial events in vascular development for gymnosperms is similar to that of Tilia.
However,
Secondary Xylem in gymnosperms is more homogenous
than that in most Angiosperms. 
The only Tracheary Elements present are Tracheids. However, in environments which have distinct seasons (Cold/Warm or Wet/Dry) large thin-walled tracheids are produced during optimal growth periods while narrower, thick-walled cells are produced under stressful conditions.
This produces "Growth Rings". Furthermore, there are more pits in the thin-walled tracheids compared to the thick-walled tracheids. Consequently, there is some degree of structure/function variation in the xylem. What does this last sentence mean???
Observe the
following from commercial slides  and fresh sections stained with Phloroglucinol.
and fresh sections stained with Phloroglucinol.
Work from the outside towards the inside of the stem!!!
Locate the Periderm, which will replace the Epidermis.
The Cortex has large resin canals. These are tube-like cavities originating as schizogenous intercellular spaces. They are lined with epithelial cells that excrete resinous material into the cavity. Many cortical parenchyma cells contain tannins.
The outermost phloem occurs just inside the resin canals. The sieve cells in the outer part of the phloem are crushed. The functional phloem occurs next to the vascular cambium.
The secondary xylem stains red due to the presence of lignin. It is fairly homogenous in appearance, but differences in diameter and wall thickness produce alternating growth rings.
The pith occupies the center of the stem.
Secondary Xylem and Phloem of Podocarpus
View cross sections stained with Phloroglucinol
The vascular tissues are similar to what you saw with Pinus.
However, the secondary phloem contains fibers. Why is this noteworthy?
|
 |
Secondary Phloem and Xylem of Podocarpus stained with Phloroglucinol |
|
![]()






