| Branching | Stele | Leaves | Roots | Buds | Stele | |
| Sexual Repro. | Micro gameto genesis |
Mega gameto genesis |
Fert. | Germination | Roots Sec. Growth | |
Over 50 Genera
Over 500 Species
Conifers have a
world-wide distribution and range from the subarctic to the subtropics.They form a
significant or dominant component of ecosystems which range from  N. Maine to S. Mississippi, and from S. California to N. Washington
(USA). They are the dominant plants in the N. Boreal Forest which runs from Subarctic
Alaska, & Canada through Scandanavia & Russia. They are also present in the S.
Hemisphere. Araucaria (Norfolk Island Pine) is native to the S. Hemisphere as are
several other species.
N. Maine to S. Mississippi, and from S. California to N. Washington
(USA). They are the dominant plants in the N. Boreal Forest which runs from Subarctic
Alaska, & Canada through Scandanavia & Russia. They are also present in the S.
Hemisphere. Araucaria (Norfolk Island Pine) is native to the S. Hemisphere as are
several other species.
Araucaria
is very invasive in Hawaii!
Conifers have adaptations which allow them to grow in stressful environments. They can survive drought, high winds, searing heat and numbing subzero cold. All of these may be found in the same location. For instance Junipers and Ponderosa Pine, growing east of the Rockies are exposed to all of the above.
Some
plants like Cypress can even grow in hydric environments that occur in the swamps
of Florida or Louisiana.
These are perennial shrubs and trees. Sequoias, for instance, are the largest organisms on the planet. They are also among the oldest in terms of longevity. Pinus longavea reputedly can live for 5,000 yrs.

Conifers have multicellular apical meristems which are similar to angiosperms.
Leaf Initiation and distribution are similar to angiosperms, and the stems show some internodal elongation.
|
|
| Conifer Shoot Apical Meristem - Note that it is Multicellular | Conifer Buds - Note the Waxy coating |
Branching generally
occurs at the nodes  and is axillary. Many conifers have
strong Apical Dominance. This results in a prominent central axis (monopodial) with
proportionately shorter branches from the bottom to the top. This yields a conical
growth form.
and is axillary. Many conifers have
strong Apical Dominance. This results in a prominent central axis (monopodial) with
proportionately shorter branches from the bottom to the top. This yields a conical
growth form.
Conifers produce weather-tight buds. These are composed of small, sclerotic leaves called bud-scales which secrete wax. They enclose vegetative and reproductive apices. The buds protect primordial structures which would otherwise perish.
Dramatic growth occurs in the spring when the dormant apices break through the bud-scales and elongate rapidly.
|
|
Mature branch of Abies (True Fir) - Note the leaves (nodes) and internodes |
Flush of new growth from a Pinus stem tip |
![]()

Link to Secondary Growth Web Pages
Seedlings have a Eustele with discrete vascular bundles. However, the vascular cambium quickly unites the bundles to produce an ectophloic siphonostele which is the mature form.
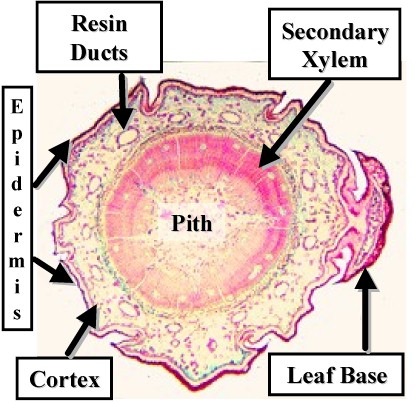 |
Cross Section of a Pine Twig - Note the Leaf Bases |

Secondary xylem is composed entirely of tracheids and ray parenchyma. Some tracheids have thicker walls and are more important for physical support than for water transport.
The rays are uniseriate and there is no axial parenchyma in the xylem.
Accumulated secondary xylem is called wood. Conifers produce a light uniform wood which can be very strong.
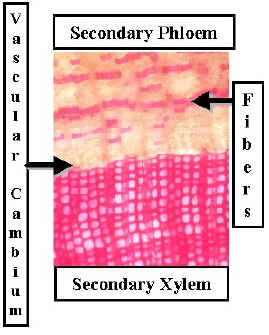
Secondary Phloem is composed of sieve cells & parenchyma. Some species produce secondary phloem fibers and some do not.
A Periderm
develops and forms a substantial bark. Certain species like Ponderosa Pine  (Pinus ponderosa)
and Douglas fir (Pseudotsuga mertensiana) are able to survive in dry grassland communities because their bark can
withstand prairie fires. The coast Redwood (Sequoia
spempervirens) and Giant Sequoia (Sequoiadendron
giganteum) are also fire resistant due to their thick
Periderm.
(Pinus ponderosa)
and Douglas fir (Pseudotsuga mertensiana) are able to survive in dry grassland communities because their bark can
withstand prairie fires. The coast Redwood (Sequoia
spempervirens) and Giant Sequoia (Sequoiadendron
giganteum) are also fire resistant due to their thick
Periderm.
|
Bark from Araucaria (Norfolk Island Pine) - Note that it is shed in fairly large sheets. This is called "Ring Bark" |
|
Bark of Pinus - Note that it is organized into small scale-like segments. This is called "Scale Bark". |

Conifer leaves
are usually needle-like although some are scale-like and others have a flat lamina.
These range in size from a few mm. to 30 cm. There is only one centrally located vascular
bundle in the leaf. However, it is often partitioned into separate strands. The vascular
tissue is surrounded by a "transfusion tissue". This is
composed of parenchyma and tracheids. An Endodermis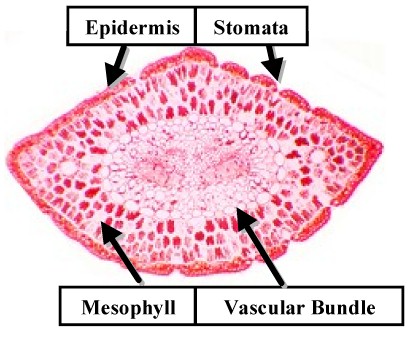 surrounds the vascular bundle.
surrounds the vascular bundle.
The photosynthetic tissue is usually not differentiated into palisade & spongy mesophyll. In Pinus the chlorenchyma cells resemble parts of a jigsaw puzzle with decidedly undulated margins. These form wafer-like layers which are separated by air spaces above and below.
A sclerefied hypodermis
frequently occurs. The epidermis has a thick cuticle
and stomata  are sunken. These traits are typical for a "xeromorphic" leaf.
Xero means dry. These adaptations help to explain why conifers can survive harsh winter
and summer conditions. These plants are commonly known as "evergreens" because
they retain their leaves during the winter or dry seasons. A great deal of energy is
invested in leaf production. Consequently, it is more efficient for leaves to function
over several years. Conifers do shed their leaves but most do so gradually over a long
time span. New leaves appear as old ones are shed. Thus, it appears that the plants never
shed their leaves. However, a stroll through a conifer-dominated forest will quickly
dispel this notion. Pine needles are allelopathic and kill understory plants.
Consequently, these are not the best plants to use for erosion control.
are sunken. These traits are typical for a "xeromorphic" leaf.
Xero means dry. These adaptations help to explain why conifers can survive harsh winter
and summer conditions. These plants are commonly known as "evergreens" because
they retain their leaves during the winter or dry seasons. A great deal of energy is
invested in leaf production. Consequently, it is more efficient for leaves to function
over several years. Conifers do shed their leaves but most do so gradually over a long
time span. New leaves appear as old ones are shed. Thus, it appears that the plants never
shed their leaves. However, a stroll through a conifer-dominated forest will quickly
dispel this notion. Pine needles are allelopathic and kill understory plants.
Consequently, these are not the best plants to use for erosion control.
Araucaria was introduced for erosion control on Oahu. It has become an invader and is slowly dominating ridges and valleys. A trip to the park at St. Lois Heights will show you how the understory and ground cover are eliminated by a dense stand of Araucaria.
I
have seen one local site of serious erosion that was planted with a pine species. This is
surely a mistake and demonstrates the need for basic biological knowledge by land
managers. 
Some conifers are completely deciduous. The leaves of Larch (Larix) turn yellow and dehisce in the autumn. New leaves appear in the spring. The dawn redwood (Sequoiadendron giganteum) is also deciduous.
|
|
Tap Root System |
Lateral Root |
Conifer Roots are very similar to angiosperm roots. There is an embryonic root which develops into a tap root system. Lateral branching occurs.
Mycorrhizae are typically associated with conifer roots and play an important role in seedling growth.
The root apex is multicellular with a prominent Root Cap. The primary tissues are arranged concentrically with xylem at the center, surrounded by phloem, and the pericycle. The endodermis and the rest of the cortex are typical. The dermal layer, however, is somewhat ambiguous as the epidermis is indistinct.
 |
|
| Typical Gymnosperm Root Apex viewed with Phase Contrast Optics | Close up of the Root Apical Meristem seen with Nomarski Optics |
Secondary meristems arise in the stele and the cortex of roots. Secondary xylem and phloem are similar to that produced in the stem, and the periderm is similar but not so pronounced.
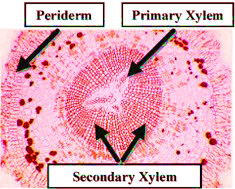 |
| Cross-Section of a gymnosperm root showing secondary growth in the stele and Periderm formation in the cortex. |
![]()
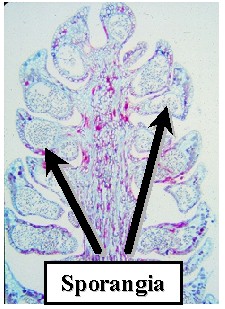
Conifers produce Microsporangia and Megasporangia in Cones.
|
Each Microsporophyll contains two Microsporangia. Each Microsporangium. produces hundreds of Microspores. These develop into Pollen Grains. Pollen grains are multicellular and constitute the Microgametophyte.
There is a Generative Cell that produces two Sperm, and a Tube Cell that produces the Pollen Tube. There are some additional cells which do not contribute directly to the reproductive process.
The mature Microgametophyte develops a pollen tube which delivers the sperm to the archegonia. At this point the sperm are released and fertilization occurs. The Microgametophyte thus has an Endosporic developmental phase and an Exosporic phase. It is protected by a thick wall when it is released from the Microsporangium. and during its flight. The latter is aided by wing-like projections. The Microgametophyte is also protected by the ovule once it enters the micropyle. However, this is still an unrefined process and billions upon billions of pollen grains are produced each year. In some cases the air turns yellow due to a blizzard of pollen grains. OK so this may be hyperbole!!!! It is notable that the sperm lack flagella and they are delivered directly to the archegonia by the pollen tube.
 |
 |
|
Microsporangiate or Pollen Cones (yellow) & Megasporangiate or Seed Cone (green) of Pinus |
Microsporangiate (Pollen) Cones of Pinus - Each unit is a sporophyll with two sporangia. |
Long section of a Pinus Microsporangiate Cone |
|
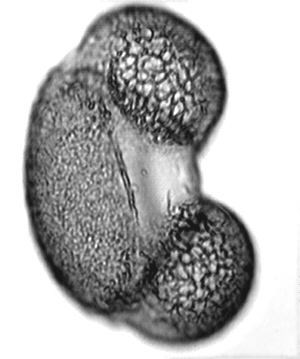 Light Microscope |
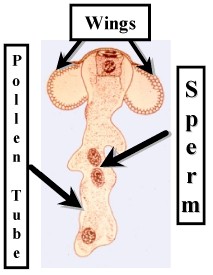 Pollen Tube |
| Conifer Microgametophyte | ||
|
|
Microsporangiate Cones of Podocarpus |
Microsporangia of Podocarpus |
Conifers are gymnosperms. Gymnosperm means naked seed. The best local example of this is Podocarpus which is a species of the Southern Hemisphere. It has strap-like leaves with a prominent "midrib" It produces Ovules at the tips of short lateral branches. The ovules are waxy and have a frosty blue appearance.
Leaves and Ovules of Podocarpus |
|
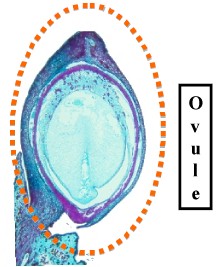 |
 |
| Long section through a Podocarpus Ovule | Mature Podocarpus Ovules with fleshy Arils |
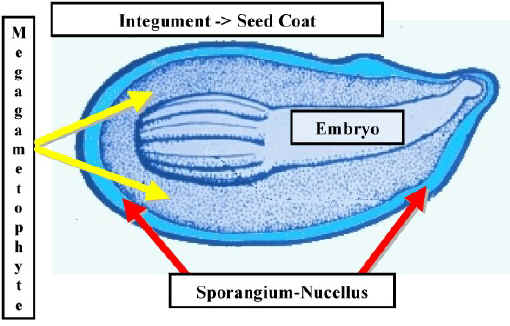
An ovule is a Megasporangium with a skin or Integument. The integument has a small opening called the Micropyle. The Micropyle secretes a drop of fluid when it is receptive. This is called a Pollination Droplet. Pollen that lands in this drop is drawn into the Micropyle as the fluid dries. After pollination, the Micropyle closes due to further growth of the ovule. Once inside the ovule, pollen grains germinate to form a pollen tube.
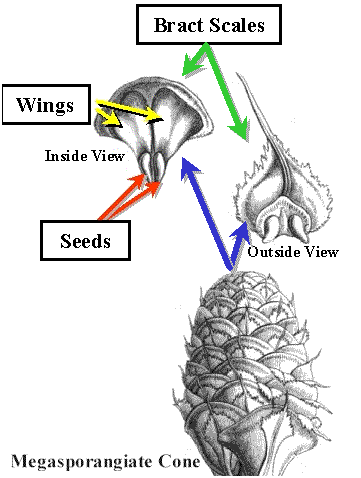
Most Conifers
produce Ovules in cones.  The unit structures in conifers are compound. They
contain a leaf bract and a highly reduced stem called a scale. This is called the bract
scale complex. However, we can think of them as "sporophylls".
Each "sporophyll" produces two Megasporangia. Meiosis
takes place in one cell within each sporangium. This results in four
haploid Megaspores. Three of these degenerate.
The remaining megaspore divides to produce a
gametophyte. The Megagametophyte
The unit structures in conifers are compound. They
contain a leaf bract and a highly reduced stem called a scale. This is called the bract
scale complex. However, we can think of them as "sporophylls".
Each "sporophyll" produces two Megasporangia. Meiosis
takes place in one cell within each sporangium. This results in four
haploid Megaspores. Three of these degenerate.
The remaining megaspore divides to produce a
gametophyte. The Megagametophyte develops within diploid tissue which is called the Nucellus.
The gametophyte grows at the expense of the Nucellus which is largely consumed in this
process. However some Nucellus remains, especially near the micropyle. Several Archegonia
develop at the micropylar end of the gametophyte. Each
of these contains a large egg.
develops within diploid tissue which is called the Nucellus.
The gametophyte grows at the expense of the Nucellus which is largely consumed in this
process. However some Nucellus remains, especially near the micropyle. Several Archegonia
develop at the micropylar end of the gametophyte. Each
of these contains a large egg.
|
|
Immature Megasporangiate Cone of Pinus |
Longitudinal section of a mature Megasporangiate cone of Pinus |
|
| Diagram of a Megasporangiate Cone and the Bract Scales |
Fertilization takes place when the pollen tubes grow through the remaining Nucellus and into the neck of the archegonia. Multiple fertilizations frequently occur and the resulting embryos compete for nutrients. Eventually one takes over and grows to maturity inside the megagametophyte.
|
Diagram of a Mature Ovule |
|
|
| Mature Ovule of Pinus - Note the large eggs which make up the greatest part of each archegonium. Note the megagametophyte which has grown inside the Nucellus. Some Nucellus is preserved, especially near the micropyle. Also note the Integument which has three distinct layers. |
|
| Conifer Embryos develop long suspensors which push the embryo proper into the megagametophyte. The embryo is linear with a shoot apical meristem at one pole and a root apical meristem at the opposite pole. Several cotyledons are located at the Distal end of the embryo. |
|
| Long Section through a Mature Conifer Embryo. The Micropyle is towards the right. The embryo is completely surrounded by the Megagametophyte. The seed coat is absent in this picture. |
|
| Mature Gymnosperm seed without its Seed-coat - Note that the embryo is surrounded by a nutritive Megagametophyte. The location of embryonic organs is not obvious in a whole mount but is inferred from microscopic data. |
The integument forms the seed coat. In many species papery wings develop from the outer integument. The wings help disperse mature seeds.
|
| Mature Seed |
|
Mature Seed & Early events in Seed Germination |
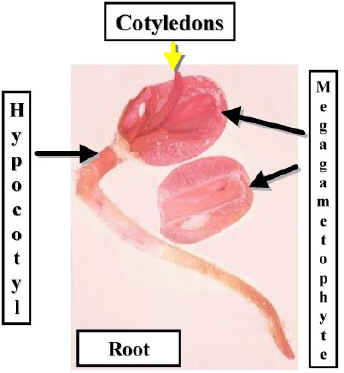
The first sign of germination is the fracturing of the seed coat and the emergence of the Radicle (root). The Hypocotyl elongates and lifts the remainder of the seed above the substrate. The Cotyledons enlarge and absorb nutrients from the Megagametophyte. However, further growth of the cotyledons results in their separation from the megagametophyte. The cotyledons complete their development and act as photosynthetic leaves.
The embryonic shoot (Epicotyl) emerges and produces the first leafy stem. This overtakes the cotyledons as the source of photosynthate.
Conifer seeds can persist in the soil for decades. They can withstand freezing and fires. Cones of lodgepole pine will not open unless they are exposed to the heat of a forest fire. Consequently, this species often forms pure stands following hot fires. They are replaced in older stands by more dominant species. However, their seeds are waiting in the soil for the next fire.
|
|
| Germinating Conifer Seedling | Diagram of a Conifer Seedling |
|
 Seedling among Moss |
 Douglas Fir Cone and Seedlings |
|
| Conifer Seedlings Germinating in Nature | |
The Seed has many obvious functional adaptations
The wings aid dispersal.
The seed is nutritive and is eaten by animals who act as dispersal agents.
The seed coat prevents the entry of pathogens and protects it from physical damage.
The strong seed coat allows seeds to be dispersed by abiotic agents without damage.
The seed contains a mature embryo which is ready to germinate (endosporic development).
The megagametophyte is a readily available food source during germination.
The seed is viable for long periods in the soil. Thus, its germination can be stretched over many years or can be triggered by specific environmental stimuli, like heat.
|
| Ponderosa Pine (Pinus ponderosa) |
|
| Giant Sequoia (Sequoiadendron gigantea) |
![]()