Botanik online 1996-2004. Die Seiten werden nicht mehr bearbeitet, sie bleiben als historisches Dokument der botanischen Wissenschaft online erhalten!
1926 untersuchte der Japaner E. KUROSAWA eine Reiskrankheit, die in Japan unter der Bezeichnung "verrückte Reiskeimlinge" bekannt war. Die Pflanzen wachsen extrem schnell, sehen spindelförmig und bleich aus und knicken wegen mangelnder Standfestigkeit leicht ab. Als Ursache für das abnorme Wachstum konnte KUROSAWA eine Substanz ausmachen, die von einem auf den Pflanzen parasitierenden Pilz (Fusarium moniliforme = Gibberella fujikuroi) ausgeschieden wird. Sie erhielt die Bezeichnung Gibberellin (GA).
Mitte der dreißiger Jahre wurde sie von japanischen Wissenschaftlern aus Tokio (YABUTA und SUMIKI) isoliert und kristallisiert, geriet dann jedoch weitgehend in Vergessenheit. 1956 wurde durch C. A. WEST und B. O. PHINNEY ein Gibberellin aus Phaseolus vulgaris und anderen Pflanzen isoliert, womit gezeigt war, daß diese Stoffklasse im Pflanzenreich weit verbreitet ist. Heute kennt man über 110 verschiedene Gibberelline (GA1, GA2...GA3, GA4...GA50), die sich chemisch z.T. nur wenig, in ihrer biologischen Aktivität jedoch sehr deutlich voneinander unterscheiden.
Etwa 30 Prozent der bekannten Gibberelline sind biologisch aktiv. Vermutlich enthalten alle höheren Pflanzen mindestens ein, in der Regel wohl mehrere aktive und inaktive Formen in unterschiedlichen Konzentrationen in den einzelnen Geweben.
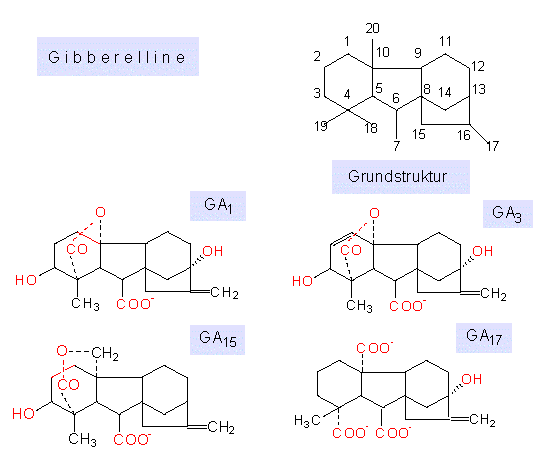
Gibberelline sind Diterpene, sie enthalten vier Isopreneinheiten, die zu einem Viererringsystem verbunden sind. Man unterscheidet zwischen den Gibberellinen mit 19 und solchen mit 20 C-Atomen. Das 20. C-Atom ist nicht Bestandteil eines der vier Ringe, sondern Teil einer Seitenkette (CH3 in GA12, CH2OH in GA15, CHO in GAl9 oder COOH in GA28). An dieser Aufzählung und den Beispielen in der Abbildung wird deutlich, in welcher Hinsicht sich die einzelnen Molekülstrukturen voneinander unterscheiden.
Reich an Gibberellinen sind junge, wachsende meristematische Achsengewebe, apikale Wurzelzellen, junge Früchte sowie unreife und keimende Samen. In Sonnenblumen ist der Gibberellingehalt in den jungen Blättern und in den oberen Internodien am höchsten, er sinkt in Richtung der basalwärts sitzenden Blätter und der basalen Internodien stetig ab.
Der Gibberellingehalt ist mit der Wachstumsgeschwindigkeit eines Gewebes korreliert. Außer im Sproßteil wurden auch in den Wurzelspitzen recht hohe Konzentrationen gemessen. Einerseits lag der Gedanke nahe, daß sie dortselbst gebildet werden, andererseits gibt es Hinweise darauf, daß dort lediglich eine Umwandlung stattfindet, d.h., daß ein Gibberellin vom Sproß in die Wurzel verfrachtet wird und dort in ein anderes umgesetzt wird. Das dabei entstehende Produkt wird via Xylem in den Sproß zurückbefördert. Sowohl in Xylem- als auch in Phloem-Exsudaten sind Gibberelline nachgewiesen worden, woraus ersichtlich ist, wie der Verteilungsmechanismus für Hormone dieser Klasse in der Pflanze funktioniert. Darüber hinaus gibt es sichere Hinweise darauf, daß es in den Geweben selbst einen Transport von Zelle zu Zelle gibt (symplastischer Transport).
Biologische Aktivität: Besonders eindrucksvoll kann die Wirkung eines Gibberellins (GA3) auf Mutanten von Phaseolus vulgaris demonstriert werden, die sich aufgrund eines genetischen Defekts durch Zwergwuchs auszeichnen. Nach Gibberellinbehandlung entstehen Pflanzen, die genauso groß wie die Kontrollpflanzen (ohne den genetischen Defekt) sind (PHINNEY, 1956). Das Ergebnis legt den Verdacht nahe, daß Zwergwuchs auf einem Defekt in der Biosynthese des betreffenden Gibberellins beruht. Vergleichbare Ergebnisse sind an Zwergmutanten anderer Kulturpflanzen erzielt worden. Durch die Gibberellinwirkung wird vornehmlich das Streckungs- und nicht das Teilungswachstum gefördert. Gibberelline stimulieren Pollenkeimung und das Auswachsen von Pollenschläuchen. Sie induzieren die Entwicklung parthenokarper Früchte, z.B. bei Äpfeln, Kürbissen und Auberginen. Durch gleichzeitige Applikation von Gibberellin und Auxin können bei einigen Arten Früchte erzeugt werden, die doppelt so groß wie die normalen sind.
Blütenbildung wird bei einer Anzahl von Pflanzenarten durch externe Faktoren, vor allem durch Licht (Kurztag- und Langtagpflanzen) oder niedrige Temperaturen (Vernalisation) gesteuert. Wenn bestimmte Langtagpflanzen oder Pflanzen, die einer Vernalisation bedürfen (Hyoscyamus, Daucus, Crepis, Silene) unter Kurztagbedingungen und/oder ohne Kältereiz gehalten werden, unterbleibt die Blütenbildung; doch nach Zusatz von GA3 blühen sie auch ohne die sonst nötigen externen Signale. Dennoch besagt diese Aussage noch nicht, daß die Gibberelline an der Blütenbildung selbst beteiligt sind, vielmehr geht bei den genannten Arten der Blütenbildung eine Verlängerung der Sproßachse voraus, und es sieht ganz so aus, als würden die Gibberelline vornehmlich diesen Entwicklungsschritt fördern. Bei den meisten anderen Langtagpflanzen (deren Sproßachse sich unmittelbar vor der Blütenbildung nicht streckt) sowie bei Kurztagpflanzen sind Gibberelline in bezug auf Blütenbildung bezeichnenderweise wirkungslos. Bei obligatorischen Lichtkeimern konnte gezeigt werden, daß Gibberelline auch in der Lage sind PFR zu substituieren. Gibberelline (so auch wieder das am besten untersuchte GA3) kontrollieren die Bildung und Sekretion von Hydrolasen in Getreidekörnern (z.B. denen der Gerste): Die Mobilisierung von Reservestoffen des Endosperms während der Keimung versorgt den Embryo mit Nährstoffen. Hieran wirken eine Reihe von Hydrolasen, so die alpha-Amylase, eine Protease und eine Ribonuklease mit. Sie werden während früher Keimungsstadien des Embryos in den Zellen der Aleuronschicht gebildet. Der Wirkungsmechanismus dieser Reaktion wurde im Labor von J. E. VARNER an der Washington University in St. Louis zu Beginn der siebziger Jahre weitgehend aufgeklärt.
Nach GA3-Einwirkung steigt die Konzentration an aktiver alpha-Amylase. Der Aktivitätsanstieg beruht auf Neusynthese. Wegen der leichten Testbarkeit erwies sich die alpha-Amylase als geeigneter Marker für eine hormonkontrollierte Enzymbildung in diesem System.
Die Bildung wird durch Transkriptionsinhibitoren (z.B. Aktinomycin D oder Chlororamphenicol) unterbunden. Simultan mit der Enzymsynthese ist eine deutliche Vergrößerung des Endoplasmatischen Retikulums und der Bildung von Polysomen erkennbar. Damit verbunden ist ein gesteigerter Einbau von Phospholipiden in die neu gebildeten Membranen sowie die Synthese der für die Membranbildung benötigten Enzyme.
Die Befunde weisen darauf hin, daß GA3 zunächst die Ausbildung einer Proteinsynthesemaschinerie fördert, an der in einem zweiten Schritt die Synthesen der genannten Hydrolasen stattfinden. Auch hier greift GA3 selektiv ein, denn es wurde nach GA3-Applikation eine forcierte Bildung von mRNS mit der Instruktion zur Bildung von alpha-Amylase nachgewiesen.
Allerdings muß auch mit allem Nachdruck darauf hingewiesen werden, daß die an Gerstenkeimlingen beobachteten Aktivitäten nur bei wenigen anderen Arten reproduzierbar waren, obwohl die auch anderwärts vorkommenden Differenzierungsschritte nach dem gleichen Schema wie bei der Gerste ablaufen. Es ist daher auch die Frage aufgeworfen worden, ob die GA3-Konzentrationsänderungen nicht lediglich Begleiterscheinungen des normalen Differenzierungsprozesses seien (P. HALMER, 1985).