Botanik online 1996-2004. Die Seiten werden nicht mehr bearbeitet, sie bleiben als historisches Dokument der botanischen Wissenschaft online erhalten!
Assimilationsgewebe im weitesten Sinne sind all jene, die aus chloroplastenhaltigen Zellen bestehen und daher zu Photosyntheseleistungen befähigt sind. Man findet sie überall dort, wo Pflanzen grün aussehen. Ein wichtiger Teilaspekt der Photosynthese ist der Einbau von Kohlendioxyd in organisches Material. Die dabei anfallenden Produkte bezeichnet man summarisch als Assimilate. Bekanntermaßen sind die Laubblätter höherer Pflanzen die bei weitem wichtigsten Produktionsstätten - sofern man aquatische einzellige Algen hier außer acht läßt. Blätter sind in der Regel aus drei Gewebetypen, dem Mesophyll, der Epidermis und dem Leitgewebe aufgebaut.
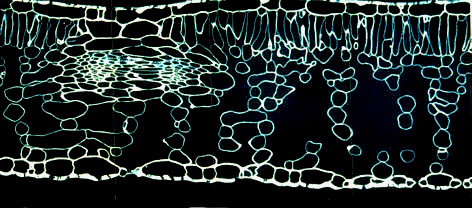
Das Mesophyll ist ein parenchymatisches Gewebe, das als das eigentliche Assimilationsgewebe angesehen werden kann. In den Blättern der meisten Farne und Phanerogamen, besonders ausgeprägt bei den Dikotyledonen und vielen Monokotyledonen, ist es in Palisaden- und Schwammparenchym untergliedert.
Bevor wir auf Einzelheiten eingehen, sei vermerkt, daß die Begriffe Mesophyll und Assimilationsgewebe nicht als Synonyme verwendet werden dürfen, denn es gibt chloroplastenfreie Blätter, die sich in ihrem Aufbau nur unwesentlich von vergleichbaren, grünen Blättern unterscheiden. Folglich enthalten sie Mesophyll, doch eine Assimilation unterbleibt; umgekehrt findet eine Assimilation z.B. auch in Sproßabschnitten statt.
Ein "typisches" Blatt ist dorsiventral gebaut. Das Palisadenparenchym liegt dabei meist an der Blattoberseite unmittelbar unter der Epidermis. Das Schwammparenchym füllt den darunterliegenden Raum aus. Es ist von einem voluminösen Interzellularsystem durchsetzt, dessen Gasraum über die Spaltöffnungen in direktem Kontakt zur Außenwelt steht.
Querschnitt durch ein Blatt von Helleborus argutifolius. Semidünnschnitt, Schnittdicke 3 µm. Die Zellwände sind mit Calcofluor white fluorochromiert.
Die hier grob skizzierte Struktur des Assimilationsgewebes wird immer wieder als ein Musterbeispiel für Selektion und Adaptation (Anpassung) zitiert. Nach dem, was wir heute über die Photosynthese wissen, muß ein effizient arbeitendes Assimilationsgewebe zumindest folgende Kriterien erfüllen:
| Es muß so strukturiert sein, daß einfallendes Licht optimal genutzt werden kann. | |
| Die Zellen bedürfen einer kontinuierlichen und ausreichenden Zufuhr an Kohlendioxyd. | |
| Alle Zellen müssen über direkten Zell-zu-Zell-Kontakt mit dem Leitungssystem verbunden sein, damit die Wasserzufuhr und ein Abtransport der Assimilate gewährleistet sind. |
Die Konstruktion des Assimilationsgewebes der Laubblätter erfüllt die genannten Bedingungen auf nahezu ideale Weise. Hinzu kommt, daß die äußere Gestalt der Blattspreite (Lamina) flächig und dünn ist und die Stellung der Blätter an Sproßhaupt- und nebenachsen den Anforderungen maximaler Lichtausnutzung entgegenkommen. Der Beschattungsgrad der Chloroplasten durch Zellen und andere Blätter wird somit auf ein Minimum reduziert. Die Versorgung der Pflanzen mit Licht, Kohlendioxyd und vor allem mit Wasser schwankt in weiten Grenzen. Daher ist es auch nicht verwunderlich, daß im Verlauf der Evolution Modifikationen des Konstruktionsplans mit erhöhtem adaptivem Wert entstanden sind, deren Ausprägung von Umwelt-/Standortbedingungen gesteuert wird.
Das Palisadenparenchym besteht aus langgestreckten, zylindrischen Zellen. Nebeneinanderliegende Zellen ähneln daher dem Bild von Pfählen, aus denen eine Palisade aufgebaut ist. Der Vergleich ist treffend, wenn man Querschnitte eines Blattes betrachtet, ist aber weniger glücklich, wenn man sich vergegenwärtigt, daß die Zellen ja nicht nur in einer Reihe, sondern auch zu einer Fläche angeordnet sind. Palisadenparenchymzellen enthalten im Vergleich zu Schwammparenchymzellen größenordnungsmäßig drei- bis fünfmal so viele Chloroplasten. Sie liegen in der Regel wandständig, da diese Anordnung eine optimale Lichtausnutzung sichert.
Bei einigen Arten sind die Zellen irregulär, gelegentlich sind sie gegabelt (Y-förmig: Armpalisadenparenchym). Solche Varianten kommen bei Farnen, Coniferen und einigen wenigen Angiospermen (z.B. bei einigen Ranunculaceen- und Caprifoliaceenarten [Beispiel: Holunder]) vor. Es gibt keine direkten verwandtschaftlichen Beziehungen zwischen diesen Arten (Gruppen), darum stellt sich die Frage: Worin liegt der Selektionswert des Armpalisadenparenchyms? Man sagt, die Zelloberfläche werde dadurch vergrößert, was vorteilhaft sei. Das mag stimmen, doch daraus ergibt sich die bislang offene Folgefrage: Warum ist diese Struktur so selten?
Die Entstehung und vor allem die Ausdifferenzierung des Palisadenparenchyms wird vielfach durch äußere Faktoren, vor allem durch Licht oder den Kohlendioxyd-Gehalt der Atmosphäre beeinflußt . Bei vielen Arten wird zwischen Sonnen- und Schattenblättern unterschieden. Sonnenblätter waren während ihrer Ontogenese einer großen Lichtmenge ausgesetzt. Es entwickelte sich ein mehrschichtiges Palisadenparenchym. Schattenblätter hingegen erhielten während der Entwicklung nur wenig Licht, das Palisadenparenchym blieb einschichtig. Die Vergrößerung des Palisadenparenchyms geht in der Regel auf Kosten der Ausdehnung des Schwammparenchyms, das bei Sonnenblättern schwächer als bei Schattenblättern ausgeprägt ist. Ein nicht minder wichtiger Faktor für den Grad der Differenzierung ist die Position eines Blattes am Sproß. Ältere, am Sproß unten stehende Blätter haben oft ein einschichtiges, junge, oben stehende, ein mehrschichtiges Palisadenparenchym. Neben der eben beschriebenen, am weitesten verbreiteten Organisationsform sind die folgenden Varianten nachgewiesen worden:
| Palisadenparenchym an der Blattunterseite. Besonders auffällig bei Schuppenblättern. Beispiel: Lebensbaum (Thuja), sowie bei den Blättern des Bärlauchs (Allium ursinum). | |
| Palisadenparenchym an beiden Blattseiten (Ober- und Unterseite). Häufig bei Pflanzen trockener Standorte (Xerophyten). Beispiel: Kompaßpflanze (Lactuca serriola). | |
| Ringförmig geschlossenes Palisadenparenchym: In zylindrisch organisierten Blättern und in Nadeln der Koniferen. |
Die Variabilität der Schwammparenchymzellen und die Ausbildung des Schwammparenchyms selbst sind noch vielgestaltiger als die des Palisadenparenchyms. Es wird meist als Durchlüftungsgewebe bezeichnet, denn es enthält eine Vielzahl untereinander verbundener Interzellularen. Das bedeutet jedoch nicht, daß der Kontakt der Palisadenparenchymzellen zu gashaltigen Interzellularen weniger stark ausgeprägt sei. Im Gegenteil: anteilsmäßig wird bei ihnen ein größerer Teil ihrer Oberfläche von Interzellularen umgeben als bei den Schwammparenchymzellen. Da sie langgestreckt und zylindrisch gebaut sind, besteht selbst bei dichtester Pakkung nur ein punktueller Kontakt zu den jeweiligen Nachbarzellen. Bei den Schwammparenchymzellen hingegen sind umfangreiche flächige Kontakte zwischen den benachbarten Zellen zu erkennen, hinzu kommen Zell-zu-Zell-Verbindungen zwischen Schwamm- und Palisadenparenchymzellen sowie zwischen Schwammparenchymzellen und den Zellen der Leitungsbahnen, wodurch ein verlustloser Wasser- und Assimilattransport erklärbar ist.
Das Interzellularsystem verursacht die Ausbildung ausgedehnter innerer Oberflächen im Blatt, wobei das Verhältnis Interzellularraum zu Gesamtzellvolumen art- und standortspezifisch ist. Als Anhaltspunkt können Verhältniswerte in der Größenordnung von 70-700 :1000 genannt werden. Auf die Oberfläche bezogen ermittelte F. M. TURREL (1934) aufgrund von Messungen und Extrapolationen auf das gesamte Blattwerk eines Trompetenbaumes (Catalpa) folgende Werte:
innere Oberfläche: 5100 Quadratmeter
äußere Oberfläche: 390 Quadratmeter
Das Verhältnis der beiden Oberflächen zueinander ist ebenfalls art- und standortspezifisch. Der Quotient ist bei Schattenblättern niedrig, bei Sonnenblättern hoch.
Die Untergliederung in die beiden Parenchymtypen (Schwamm- und Palisadenparenchym) erfolgte im Verlauf der Evolution schrittweise. Beide gehen aus dem gleichen Grundgewebe hervor, und wie zahlreiche Übergänge belegen, sind die typischen Palisaden- und Schwammparenchymzellen lediglich als Extremwerte (Endglieder) einer abgeschlossenen Differenzierung zu betrachten. Von wenigen Ausnahmen abgesehen, sind bei den Algen alle Zellen zur Photosynthese befähigt. Blätter einfacher Laubmoose sind ein-, selten zweischichtig. Hier entfällt sogar die Untergliederung in Epidermis und Mesophyll.
Bei manchen anderen Moosen, so bei den Polytrichum-Arten (Laubmoos) und Riccia-Arten (Lebermoos) befinden sich auf der Blattoberseite lamellenartig aussehende Rippen, die aus mehreren übereinanderliegenden Zellreihen bestehen und auf Assimilation spezialisiert sind. Andere Lebermoose (typisches Beispiel: Marchantia) verfügen über einen mehrschichtigen, dem Boden direkt aufliegenden Thallus. Über einer Schicht großlumiger Parenchymzellen liegt ein Assimilationsgewebe, das aus zahlreichen kleinen, rundlichen, zu Gruppen vereinten Zellen und voluminösen, zusammenhängenden Luftkammern besteht.
Bei Lebermoosen sind weder Spaltöffnungen noch Schließmechanismen vorhanden. Stattdessen ist die oberseitige Epidermis von charakteristisch gebauten - nicht verschließbaren - Atemöffnungen durchsetzt.
Bei vielen Wasser- und Sumpfpflanzen tritt anstelle der Gliederung in Palisaden- und Schwammparenchym ein homogen strukturiertes Aerenchym auf, das sich durch überproportional große Lufträume (Interzellularen) auszeichnet. In den Blättern der Wasserpest (Elodea canadensis) ist das Assimilationsgewebe auf zwei Zellagen reduziert. Auch hier kann man weder von Epidermis noch von Mesophyll sprechen.
Die Leitbündel nahezu aller Gefäßpflanzen sind von der Bündelscheide, einer mehr oder weniger umfangreichen Parenchymschicht, umgeben. Meist enthalten deren Zellen weniger Chloroplasten als das übrige Mesophyll. Oft ist in ihnen Stärke gespeichert, so daß sich auch der Begriff Stärkescheide eingebürgert hat.
Bei vielen - vor allem tropischen - Gramineen (Mais, Zuckerrohr u.a.) sind die Zellen der Bündelscheiden von einem Kranz chloroplastenhaltiger Mesophyllzellen umgeben. Beide Zellschichten umgeben die Leitbündel - im Querschnitt gesehen - wie zwei konzentrische Ringe. Wie sich in den letzten Jahren herausstellte, ist dieser Bauplan für die C4-Pflanzen charakteristisch. Dabei handelt es sich um eine Gruppe von Arten aus den verschiedensten Familien der Mono- und Dikotyledonen, bei denen ein Einbau von Kohlendioxyd in eine organische Säure der eigentlichen Photosynthese vorgeschaltet ist. Pflanzen ohne diesen Zusatzweg (der einer vorübergehenden Kohlendioxyd-Speicherung dient) werden vom Standpunkt der Photosyntheseforschung aus als C3-Pflanzen bezeichnet. Selbst innerhalb einzelner Gattungen kann eine Art als C3-, eine zweite als C4-Pflanze charakterisiert werden. Das bekannteste Beispiel hierfür sind die beiden Atriplex-Arten A. patula (C3) und A. rosea (C4).
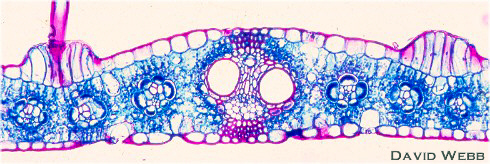
Querschnitt durch ein Monokotylenblatt (Zea mays) mit deutlich erkennbarer Kranzanatomie.
© David T. WEBB - University of Hawaii at Manoa: BOT311: Form & Function in Algae & Plants
Es gibt sogar Arten, deren untere Blätter nach dem einen, die oberen nach dem anderen Schema organisiert sind und dementsprechend einen unterschiedlichen Effizienzgrad der Photosynthese aufweisen. Es handelt es sich dabei meist um Sukkulenten (dickfleischige Pflanzen) trockener Standorte (CAM-Pflanzen). Zur Vermeidung von Transpirationsverlusten sind die Spaltöffnungen tagsüber geschlossen. Kohlendioxyd kann daher nur nachts aufgenommen und akkumuliert werden. Die Bindung erfolgt wie bei den C4-Pflanzen; die organische Säure (meist Äpfelsäure/Malat) wird in den Vakuolen großvolumiger, weitgehend chloroplastenfreier Parenchymzellen gespeichert. Tagsüber wird das Kohlendioxyd wieder abgespalten, es diffundiert von dort in die Mesophyllzellen und wird in den Photosyntheseprozeß eingeschleust.
Blätter entwickeln sich aus Blattanlagen, die ihrerseits aus kleinen seitlichen Ausstülpungen des Sprosses hervorgehen. Durch bevorzugte Teilungsrichtungen verursacht, entsteht eine flächige Struktur. Die zentral liegenden Zellen (das Plattenmeristem) sind u.a. als die eigentlichen Vorläufer der Mesophyllzellen anzusehen. Die Ausdifferenzierung in typische Palisaden- und Schwammparenchymzellen erfolgt durch ungleiche Wachstumsraten der verschiedenen Zellschichten im sich entwickelnden Blatt. Zellteilungs- und Zellstreckungsaktivität sind in den einzelnen Lagen zeitlich gegeneinander versetzt. Hinzu kommt, daß sich die Zellen in unterschiedliche Richtungen ausbreiten. Zur Veranschaulichung ein Beispiel: Zellen des Palisadenparenchyms strecken sich senkrecht zur Ebene des Blattes, Zellen des Leitgewebes, das während der Blattentwicklung ebenfalls angelegt wird, parallel zur Blattebene. Solange Teilung und Streckung in zwei Geweben (z.B. der Epidermis und dem Mesophyll) aufeinander abgeglichen sind, bleiben die Relationen der Zellen zueinander gleich. Zu einem relativ frühen Zeitpunkt jedoch beginnen die Epidermiszellen der Oberseite sich schneller als die noch nicht voll ausdifferenzierten Palisadenparenchymzellen zu teilen. Folglich entstehen durch Auseinanderweichen der letzteren große Interzellularräume. Sie werden zum Teil wieder aufgefüllt, weil sich die Palisadenparenchymzellen zu einem späteren Zeitpunkt antiklin teilen und damit ihre Zahl vermehren.
Die Wechselwirkungen zwischen Ausdifferenzierung der unteren Epidermis und dem Schwammparenchym sind variabler gestaltet. Je nach Art wird erst die Teilungsaktivität der Epidermiszellen oder erst die der Schwammparenchymzellen eingestellt. Zellteilungs- und Zellstreckungsaktivität sind im Schwammparenchym zeitlich voneinander getrennt. Die unterschiedliche Wachstumsgeschwindigkeit in den verschiedenen Teilen des sich entwickelnden Blattes führt dazu, daß sich die Zuwachszonen wellenförmig über das Blatt ausdehnen. Erst kürzlich konnten diese Wachstumsbewegungen in einem Zeitrafferfilm dokumentiert werden, in dem eindrucksvoll zu sehen ist, daß das Wachstum und die Ausdehnung eines Buchenblattes mit intensiven Wellenbewegungen einhergeht (W. KAUSCH, Universität Bonn, 1986, Herstellung und Verleih: Institut für den Wissenschaftlichen Film, Göttingen).