Bacteriorhodopsin
aus Halobacterium halobium
Bakterienrhodopsin (bR) ist eine Protonenpumpe aus der Cytoplasmamembran von Purpurbakterien. Die hier vorgestellten Daten stammen aus einer Reihe verschiedener Experimente, u.a. der Analyse zweidimensionaler und dreidimensionaler Kristalle. bR wird als Protein von 262 Aminosäuren synthetisiert, von denen ein Propeptid von 13 Aminosäuren abgespaltet wird. In dem in die Membran integrierten Protein konnte für die Aminosäuren 7 - 227 ein Modell der Atomkoordinaten erstellt werden.
bR ist ein primäres Transportsystem, Lichtenergie der Wellenlänge 568 nm wird für die Beförderung von Protonen gegen einen Konzentrationsgradienten benutzt. Sowohl der Kofaktor Retinal als auch das Protein erfahren bei der Reaktion Konformationsänderungen, die sich u.a. in unterschiedlichen Absorptionsmaxima des Komplexes äußern. Die Zwischenstufen im Reaktionszyklus sind:
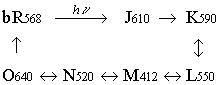
Die hier benutzten Datenbank-Atomkoordinaten beschreiben nur den Grundzustand bR568.
Die Aminosäurekette des bR ist zu sieben transmembran-Helices gefaltet, wobei das Aminoende in das Periplasma ragt und das Carboxyende in das Cytoplasma
